The Feathered Revolution: How Dinosaurs Became Birds

The evolutionary tale of birds is full of twists, turns and discoveries that continue to rewrite the history books. Paul Willis leads us through the maze.
After agonising over it for 20 years, Darwin finally published his theory of evolution, On The Origin Of Species, in 1859. But it was Darwin’s ‘bulldog’, Thomas Henry Huxley, who braced to defend the theory. To do so, Huxley desperately needed fossil evidence of a ‘missing link’ to show how animals had transitioned from one species to another – evidence that Darwin himself admitted was sadly lacking.
Just two years later in Bavaria, the Jurassic-aged limestone deposits yielded a near-perfect fossil of Archaeopteryx. It had blade-like serrated teeth and many other features across the skeleton and skull that showed it was a carnivorous dinosaur. But the crow-sized specimen was covered in the impressions of bird-like feathers. For Huxley, this was the transitional form he was seeking: a dinosaur on its way to becoming a bird. The Germans referred to it as Urvogel, the first bird (its scientific name is derived from the Greek words, ancient feathers).
It was a coup for Huxley. It was also the beginning of the feather revolution.
Archaeopteryx’s halo of feathery impressions may have been a 19th century game changer, but feathers were only just starting to overturn the evolutionary paradigm of the day.
Fast forward 137 years, and new discoveries of fossils with quills are continuing to rewrite the textbooks, not just on bird origins but across the entire dinosaur family tree.
The attempt by palaeontologists to retrace the path of bird evolution makes for a rolicking tale full of sudden twists and turns. For starters, Archaeopteryx did not settle the matter of bird origins. In the early years of the 20th century, Huxley’s proposition that birds descended from carnivorous dinosaurs, specifically the suborder known as theropods, fell out of favour.
One problem with the theory was that the skeletons of theropods were missing a crucial part of bird anatomy – the wishbone (furcula). It acts like a spring to assist flight and is made from the fusion of two collarbones. So for the first half of the 20th century, the search was on for a non-dinosaur ancestor to the birds.
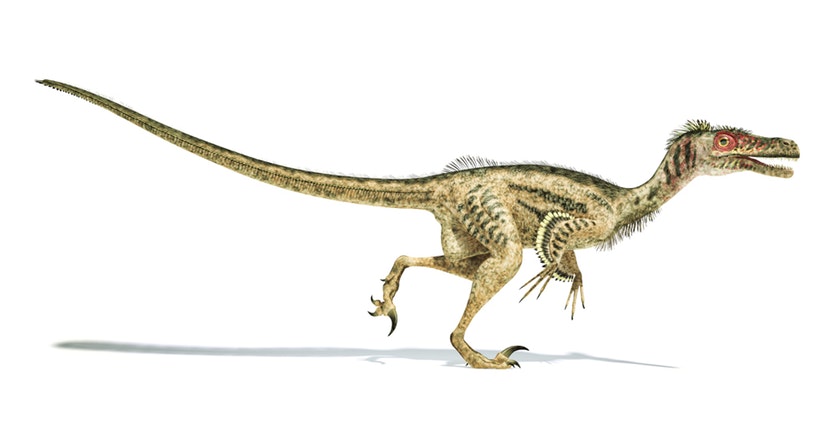
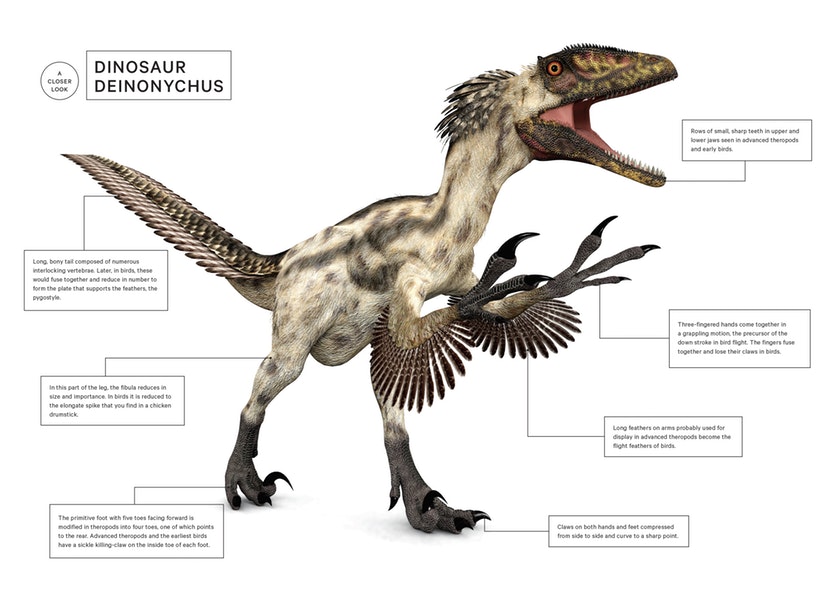
The next twist in the tale of bird evolution was added by American palaeontologist John Ostrom. He resurrected Huxley’s theory by showing numerous similarities between the skeletons of Deinonychus, a theropod from the Montana Badlands, and Archaeopteryx. Ostrom was able to show that Deinonychus and other theropods did actually have a furcula; it had previously been mistaken for an extra pair of ventral ribs. Even more exciting was the fact that this fusion of the two collarbones had clearly occurred in the theropods well in advance of the evolution of flight capability.
But it was feathers that provided the final incontrovertible evidence that birds evolved from dinosaurs. In the early 1990’s researchers began recovering extraordinary fossils of a wide range of creatures from the late Jurassic and early Cretaceous periods (150 million to 100 million years ago).
They came from Liaoning province in northern China, and, unlike most fossils, their soft tissues were still preserved.
In 1996 the Liaoning deposits surrendered their first feathered dinosaur, the 1.5 metre long theropod dinosaur, Sinosauropteryx. While it had no wings, it was covered in a feathery fuzz.
Since then, a spectacular array of small, feathered dinosaurs have been recovered from Liaoning and a few other sites around the world, which plot every conceivable evolutionary step from small fuzz-covered, meat-eating theropods through to fully feathered and winged birds.
This transition is not so much a linear path as a dense maze, with many paths leading to dead ends.

It’s also clear now that Archaeopteryx was not the first bird-like dinosaur. That honour (at least for the time being) goes to Xiaotingia zhengi. Unearthed from Liaoning province in 2011, this feathered fossil is estimated to be at least five million years older than the dozen odd specimens of Archaeopteryx found to date. Xiaotingia displayed several primitive features, such as a sickle-shaped killing claw on the second toe of each foot – the type seen on Deinonychus and the villainous Velociraptor depicted in the movie Jurassic Park. (The movie version is actually about as big as Deinonychus while the real Velociraptor is about half that size).
Xiaotingia not only had fuzz, it had a set of feathered wings capable of gliding flight, probably an intermediate step toward fully powered flight.
While Xiaotingia had a single set of wings, other gliding protobirds like the slightly younger Microraptor developed two sets. Effectively they would have looked and functioned like the wings of a biplane, with the rear set held below the front pair to avoid turbulence. A number of other small dinosaur protobirds also had elongated feathers on their hind limbs; however, in most cases, these appear to be ornamental rather than flight worthy.
But it seems dinosaur flight experiments were not restricted to feathers. In 2015 came news of a very weird theropod named Yi qi that was only distantly related to the protobirds. Found in Hebei province, which borders Liaoning, Yi is around 160 million years old, the size of a small bantam and estimated to have weighed around 380 grams.
What was so weird about this little fella was only apparent because of the extraordinary preservation of skin and feathers alongside the front end of the skeleton. Similar to its close relatives, Yi had an elongated third finger as well as a bony strut extending from the wrist.
But the beautifully preserved specimen also shows a flap of skin stretching between that long finger, the bony strut and the side of the body. Rather than using its feathers for flight, Yi was gliding on bat-like wings.
With all this aerial experimentation going on, the question remains: why were all these dinosaurs seemingly hell bent on learning to fly, with feathers or without?
The short answer seems to be that small theropods had already taken to the trees. They probably did so to avoid larger predators and to source meaty prey such as insects and small vertebrates nestled among the branches. Their curved, needle-point theropod claws on both the fingers and toes weren’t just great for grabbing prey; they also made excellent grappling hooks for climbing.
Once they were arboreal (tree-dwellers), the ability to glide from tree to tree like flying lizards and possums do today could have allowed them to extend their range and chase other gliders while staying clear of dangers on the ground. When it came to evolving true aeronautical ability, they already had a head-start thanks to a suite of features they had inherited from their theropod ancestors.
For starters, they were lightweight. All theropods had hollow but strong bones, an adaptation their ancestors developed to be faster at hunting prey.
Being lightweight helped with gliding, but progressing to powered flight depended on another inherited feature. The shoulder sockets of theropods had rotated so that instead of swinging the arms backward and forward for walking, the arms could come together in front of the animal in a grasping motion to capture prey. This grappling action also appears to be assisted by the furcula acting in a spring-like manner, linking the muscles in the chest to the actions of the arms. Any gliding protobird using this grasping action would be performing the first down stroke of powered flight.
A further modification was the development of pockets extending from the lungs, which not only increased overall lung capacity but led to a one-way breathing system where air circulates continually through the lungs instead of being pumped in and out the way humans breathe. This breathing system, a characteristic of all modern birds, was also very useful for a pursuit predator.
Feathers, which evolved alongside these other avian characteristics, would complete the journey towards modern birds. Beginning as simple downy filaments, they had probably originated to keep theropods warm – evidence supporting the theory that at least some dinosaurs were ‘warm-blooded’. They may also have served as ornaments. Festooned with arm feathers, smaller theropods’ plumage would have given them a degree of lift to chase flying insects.
This set of features – small with lightweight bones, rotated shoulder sockets with a furcula, turbocharged breathing systems and feathers – developed in theropods long before they took to the air and set the scene for protobirds.
Fossil quills have already forced multiple revisions of textbooks. And still, a finding just last year showed that feathers are not done with their revolutionary work.
For 130 years, we thought we understood the broad architecture of the dinosaur family tree. British palaeontologist Harry Seeley pointed out in 1887 that dinosaurs could be divided into two groups based on whether their hips were lizard-like (where the pubis points forward) or bird-like (where the pubis points back). Confusingly, it was members of the lizard-hipped rather than bird-hipped variety that gave rise to birds.
The lizard-hipped saurischians were in turn divided between the long-necked plant-eating sauropods, such as Brachiosaurus, and the meat-eating theropods.
The bird-hipped ornithischians included a huge variety of plant-eating dinosaurs that could be divided into three smaller groups: the armoured dinosaurs including Stegosaurus, the bird-footed ornithopods such as Iguanodon and the horned dinosaurs like Triceratops.

Bottom line: Brachiosaurus and Tyrannosaurus were relatively close cousins. Iguanodon, Stegosaurus and Triceratops were more distantly related.
Palaeontologists were quite happy with this binary arrangement until 2017, when another British palaeontologist, Matthew Baron from Cambridge University, completely redrew the family tree.
Baron looked at 74 species of exceedingly rare early dinosaurs from the first half of the Age of Dinosaurs. By analysing a very large set of characters from all over the skeletons, he was able to tease out how the early branches divided right down at the base of the tree.
His first finding pushed back the origin of dinosaurs by around five million years to about 247 million years ago. The second completely rewrote dinosaurian prehistory. Instead of a neat, early split between the lizard-hipped and bird-hipped branches, Baron found an even earlier split that placed the lizard-hipped theropods onto the same branch as the bird-hipped group.
So now Tyrannosaurus is nestled in with Triceratops (see figure opposite).
This new and, at present, controversial arrangement of dinosaur relationships lay hidden in a soup of confusing lumps and bumps on bones; there was no single feature that you could point to and say “dinosaur X belongs on this branch or that”.
Except, perhaps, feathers.
Feathers and their hairy antecedents have been found on many theropod dinosaurs and also on several different bird-hipped dinosaurs. But they have never been found among the sauropods. So the new group of dinosaurs consisting of the theropods and the bird-hipped dinosaurs (which has been named the ornithoscelidans) may be defined by the presence of feathers or feather-like structures covering some portion of the animal.
So feathers may be telling us a lot more about the structure of the dinosaur’s family tree than simply “here are the birds”. They may actually be the defining feature of one of the most fundamental splits in the dinosaur tree that occurred very early in their evolution.
The revolution is still underway and many of the preliminary conclusions presented here are far from settled and confirmed. There is still a lot of work to be done, more species to find and classify.
Long live the revolution of the feather!

Source: https://cosmosmagazine.com







