Hypacrosaurus

Hypacrosaurus was a genus of duckbill dinosaur similar in appearance to Corythosaurus. Like Corythosaurus, it had a tall, hollow rounded crest, although not as large and straight. It is known from the remains of two species that spanned 75 to 67 million years ago, in the Late Cretaceous of Alberta, Canada, and Montana, United States, and is the latest hollow-crested duckbill known from good remains in North America. It was an obscure genus until the discovery in the 1990s of nests, eggs, and hatchlings belonging to H. stebingeri.
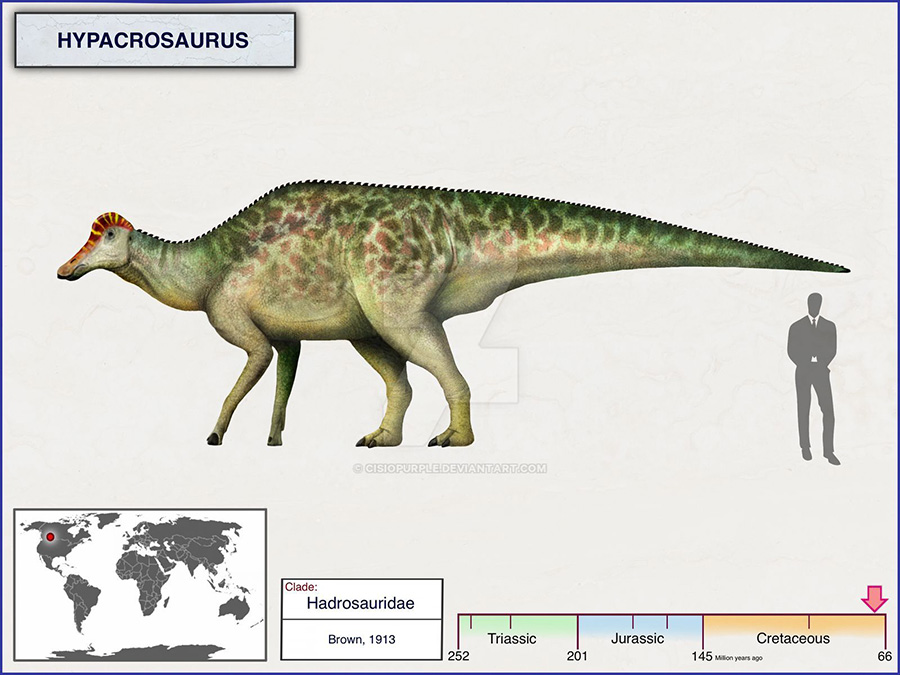
Hypacrosaurus is most easily distinguished from other hollow-crested duckbills (lambeosaurines) by its tall neural spines and the form of its crest. The neural spines, which project from the top of the vertebrae, are 5 to 7 times the height of the body of their respective vertebrae in the back, which would have given it a tall back in profile. The skull's hollow crest is like that of Corythosaurus, but is more pointed along its top, not as tall, wider side to side, and has a small bony point at the rear. Unlike other lambeosaurines, the passages for the airways do not form an S-curve in the crest (at least not in H. altispinus). The animal is estimated to have been around 9.1 meters (30 feet) long, and to have weighed up to 4.0 tonnes (4.4 tons). As with most duckbills, its skeleton is otherwise not particularly remarkable, although some pelvic details are distinctive. Like other duckbills, it was a bipedal/quadrupedal herbivore. The two known species, H. altispinus and H. stebingeri, are not differentiated in the typical method, of unique characteristics, as H. stebingeri was described as transitional between the earlier Lambeosaurus and later Hypacrosaurus. Photographs of an adult H. stebingeri skull show an animal that looks very similar to H. altispinus.
The type remains of Hypacrosaurus remains were collected in 1910 by Barnum Brown for the American Museum of Natural History. The remains, a partial postcranial skeleton consisting of several vertebrae and a partial pelvis (AMNH 5204), came from along the Red Deer River near Tolman Ferry, Alberta, Canada, from rocks of what is now known as the Horseshoe Canyon Formation (early Maastrichtian, Upper Cretaceous). Brown described these remains, in combination with other postcranial bones, in 1913 as a new genus that he considered to be like Saurolophus. No skull was known at this time, but two skulls were soon discovered and described.

Hypacrosaurus was a lambeosaurine hadrosaurid, and has been recognized as such since the description of its skull. Within the Lambeosaurinae, it is closest to Lambeosaurus and Corythosaurus, with Jack Horner and Phil Currie (1994) suggesting that H. stebingeri is transitional between Lambeosaurus and H. altispinus, and Michael K. Brett-Surman (1989) suggesting that Hypacrosaurus and Corythosaurus are the same genus. These genera, particularly Corythosaurus and Hypacrosaurus, are regarded as the "helmeted" or "hooded" branch of the lambeosaurines, and the clade they form is sometimes informally designated Lambeosaurini. Although Suzuki et al.'s 2004 redescription of Nipponosaurus found a close relationship between Nipponosaurus and Hypacrosaurus stebingeri, indicating that Hypacrosaurus may be paraphyletic, this was rejected in a later, more comprehensive reanalysis of lambeosaurines, which found the two species of Hypacrosaurus to form a clade without Nipponosaurus, with Corythosaurus and Olorotitan being the closest relatives.

As a hadrosaurid, Hypacrosaurus would have been a bipedal/quadrupedal herbivore, eating a variety of plants. Its skull permitted a grinding motion analogous to chewing, and its teeth were continually replacing and packed into dental batteries that contained hundreds of teeth, only a relative handful of which were in use at any time. Plant material would have been cropped by its broad beak, and held in the jaws by a cheek-like organ. Its feeding range would have extended from the ground to about 4 m (13 ft) above.
Crest functions
The hollow crest of Hypacrosaurus most likely had social functions, such as a visual signal allowing individuals to identify sex or species, and providing a resonating chamber for making noises. The crest and its associated nasal passages have also figured in the debate about dinosaur endothermy, specifically in discussions about nasal turbinates.

Turbinates are thin bones or cartilages that come in two types, with two functions. Nasal olfactory turbinates are found in all living tetrapods and function in smell. Respiratory turbinates function to prevent water loss through evaporation and are found only in birds and mammals, modern endotherms (warm-blooded animals) who could lose a great deal of water while breathing because they breathe more often than comparably sized ectotherms (cold-blooded animals) to support their higher metabolism. Ruben and others in 1996 concluded that respiratory turbinates were probably not present in Nanotyrannus, Ornithomimus or Hypacrosaurus based on CT scanning, thus there was no evidence that those animals were warm-blooded.

The discovery of tooth marks in the fibula of a Hypacrosaurus specimen inflicted by a bite from the teeth of a tyrannosaurid indicated that this, and other hadrosaurids were either preyed upon or scavenged by large theropod dinosaurs during the Late Cretaceous period.
Examining the oxygen-isotope ratio from the bones from different parts of an extinct animal's body should indicate which thermoregulation mode an animal used during its lifetime. An endothermic (warm-blooded) animal should maintain a very similar body temperature throughout its entire body (which is called homeothermy) and therefore there should be little variation in the oxygen-isotope ratio when measured in different bones. Alternatively, the oxygen-isotope ratio differs considerably when measured throughout the body of an organism with an ectothermic (cold-blooded) physiology. Oxygen-isotope ratios calculated for Hypacrosaurus suggesting that the ratios varied little, indicating that Hypacrosaurus was a homeotherm, and likely was endothermic. This is in contrast to the Ruben et al. (1996) finding that Hypacrosaurus was not warm-blooded, which was based on the absence of nasal turbinates (see Crest functions subsection, above).
Source: https://en.wikipedia.org/







